Implantation (embryology)
Implantation in embryology refers to the process in which a fertilized egg (or blastocyst) adheres to the wall of the uterus or womb. This process is crucial for the initiation of pregnancy as it establishes a connection between the mother and the developing embryo.
Process[edit]
Implantation begins approximately one week after fertilization. The blastocyst, which has been moving through the fallopian tube, enters the uterus. Here, it adheres to the uterine wall in a process facilitated by various hormones and cytokines. The blastocyst then burrows into the uterine wall, embedding itself in the endometrium. This process is known as "invasion".
Role in Pregnancy[edit]
Successful implantation is a critical step in establishing a healthy pregnancy. It allows the embryo to receive nutrients and oxygen from the mother, which are essential for its growth and development. If implantation does not occur, the fertilized egg will be expelled from the body during the woman's next menstrual cycle.
Factors Influencing Implantation[edit]
Several factors can influence the success of implantation. These include the health and age of the woman's eggs, the quality of the man's sperm, and the health of the woman's uterus. Certain medical conditions, such as endometriosis or polycystic ovary syndrome (PCOS), can also affect implantation.
Complications[edit]
Complications can arise if the blastocyst implants outside the uterus, such as in the fallopian tube. This is known as an ectopic pregnancy and is a serious medical condition that requires immediate treatment.
See Also[edit]

This WikiMD article can only be edited by registered and verified editors. You can log in or register.
Implantation (embryology)[edit]
-
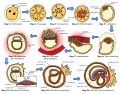 Human embryogenesis diagram
Human embryogenesis diagram -
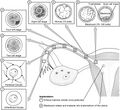 Preembryonic development
Preembryonic development -
 Implanting embryo
Implanting embryo -
 Placenta formation
Placenta formation -
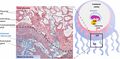 Feto-maternal interface showing uterine milk
Feto-maternal interface showing uterine milk





Sponsored Health Resource

W8MD Weight Loss, Sleep & MedSpa
Looking for physician-supervised weight loss, semaglutide, tirzepatide, or GLP-1 receptor agonist options? W8MD helps eligible patients in New York City, Brooklyn, New Jersey, Connecticut, Pennsylvania, Delaware, and greater Philadelphia with medical weight loss, sleep medicine, and long-term maintenance support.
GLP-1 specials: Affordable GLP-1 injections NYC and Philadelphia starting from $29.99/week and up for semaglutide with insurance accepted for qualifying visits, and $45/week and up for tirzepatide with insurance accepted for qualifying visits. Self-pay options start from $59.99/week and up for semaglutide and $69.99/week and up for tirzepatide.
- Medical weight loss NYC
- Affordable GLP-1 injections NYC
- Budget GLP-1 weight loss shots Philadelphia
- New Jersey medical weight loss
- NYC medical weight loss blog
- Philadelphia weight loss blog
- Sleep medicine and sleep apnea services
- W8MD MedSpa and wellness
Book a W8MD appointment · View GLP-1 specials
Paid promotional message. Eligibility, pricing, insurance coverage, medication availability, and results vary. Medical evaluation required.
")
Medical Disclaimer: WikiMD is for informational purposes only and is not a substitute for professional medical advice. Content may be inaccurate or outdated and should not be used for diagnosis or treatment. Always consult your healthcare provider for medical decisions. Verify information with trusted sources such as CDC.gov and NIH.gov. By using this site, you agree that WikiMD is not liable for any outcomes related to its content. See full disclaimer.
Credits:Most images are courtesy of Wikimedia commons, and templates, categories Wikipedia, licensed under CC BY SA or similar.
Translate page: - East Asian
中文,
日本,
한국어,
South Asian
हिन्दी,
தமிழ்,
తెలుగు,
Urdu,
ಕನ್ನಡ,
Southeast Asian
Indonesian,
Vietnamese,
Thai,
မြန်မာဘာသာ,
বাংলা
European
español,
Deutsch,
français,
Greek,
português do Brasil,
polski,
română,
русский,
Nederlands,
norsk,
svenska,
suomi,
Italian
Middle Eastern & African
عربى,
Turkish,
Persian,
Hebrew,
Afrikaans,
isiZulu,
Kiswahili,
Other
Bulgarian,
Hungarian,
Czech,
Swedish,
മലയാളം,
मराठी,
ਪੰਜਾਬੀ,
ગુજરાતી,
Portuguese,
Ukrainian
