Heart

Editor-In-Chief: Prab R Tumpati, MD
Obesity, Sleep & Internal medicine
Founder, WikiMD Wellnesspedia &
W8MD's weight loss doctor NYC
Philadelphia GLP-1 weight loss and GLP-1 clinic NYC
Muscular organ responsible for pumping blood through the body
[[File: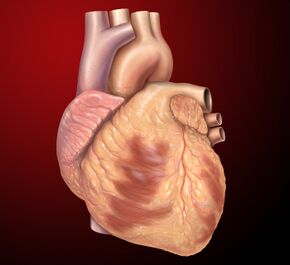 |frameless|alt=]]
|frameless|alt=]]

| Details | |
|---|---|
| Synonyms | |
| Pronunciation | |
| Carnegie stage | |
| Days | |
| Precursor | Mesoderm |
| Gives rise to | |
| Part of | |


.svg)
The heart is a hollow, muscular organ that pumps blood throughout the body's blood vessels by repeated, rhythmic contractions. It is present in all animals possessing a circulatory system, notably the vertebrate species. The term "cardiac," meaning "related to the heart," is derived from the Greek καρδιά (kardia), meaning "heart". The medical specialty dedicated to the heart, its disorders, and abnormalities is known as cardiology.
Structure[edit]
The structure and complexity of the heart can vary considerably among different animal groups.
Vertebrate heart[edit]
The vertebrate heart is primarily composed of specialized cardiac muscle tissue, connective tissue, and a lining of endothelium. Cardiac muscle is a specialized, involuntary striated muscle, uniquely adapted to sustain continuous rhythmic contractions throughout an organism’s lifespan.
Variations among animals[edit]
Cephalopods, such as octopuses and squids, possess three hearts—two "gill hearts" for pumping blood through the gills, and one systemic heart that circulates blood throughout the body. Vertebrates typically have a single heart located in the anterior portion of the body cavity, dorsal to the digestive organs.
In certain primitive vertebrates, like jawless fishes (hagfish), there is an additional heart-like structure located in the tail region, unique among vertebrates.
Human heart[edit]
In humans, the heart is a muscular organ approximately the size of a closed fist, weighing around 250–300 grams (9–11 oz) in females and 300–350 grams (11–12 oz) in males. Located within the thoracic cavity, it sits anterior to the vertebral column and posterior to the sternum, slightly left of the body's midline.
Pericardium[edit]
The human heart is enclosed within a protective double-walled sac known as the pericardium. The outer layer, called the fibrous pericardium, anchors the heart to surrounding structures and prevents over-expansion. Beneath this lies the serous pericardium, composed of two layers:
- Parietal pericardium (outer serous layer)
- Visceral pericardium (inner serous layer or epicardium, directly attached to the heart)
Between these layers lies the pericardial cavity, which contains pericardial fluid, allowing frictionless heart movements.
Layers of the heart wall[edit]
The heart wall comprises three distinct layers:
- Epicardium (outermost layer; same as the visceral pericardium)
- Myocardium (middle, muscular layer responsible for contractions)
- Endocardium (innermost layer, lining heart chambers and valves, continuous with the blood vessel endothelium)
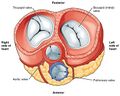
Chambers and valves[edit]
The human heart contains four chambers:
- Two superior receiving chambers: the right atrium and left atrium
- Two inferior pumping chambers: the right ventricle and left ventricle
Blood flow is controlled by four valves:
- Tricuspid valve (right atrioventricular valve, between right atrium and right ventricle)
- Pulmonary valve (between right ventricle and pulmonary artery)
- Mitral valve (left atrioventricular or bicuspid valve, between left atrium and left ventricle)
- Aortic valve (between left ventricle and the aorta)
Blood circulation[edit]
The heart supports two simultaneous circulation circuits:
Pulmonary circulation[edit]
Deoxygenated blood from systemic circulation enters the right atrium via the superior and inferior vena cavae. Blood then moves through the tricuspid valve into the right ventricle, which contracts to push blood through the pulmonary valve into the pulmonary arteries, carrying blood to the lungs for oxygenation.
Systemic circulation[edit]
Oxygen-rich blood from the lungs returns to the left atrium via the pulmonary veins. It then flows through the mitral valve into the left ventricle. Powerful contractions of the left ventricle pump blood through the aortic valve into the aorta, supplying oxygenated blood to the entire body.
Cardiac cycle[edit]
The rhythmic contraction and relaxation of heart chambers constitute the cardiac cycle, comprising two phases:
- Systole (contraction of ventricles, forcing blood into circulation)
- Diastole (relaxation phase, allowing chambers to refill with blood)
At rest, the typical adult heart beats about 72 times per minute, circulating approximately 4.7–5.7 liters of blood per minute. Over a lifespan averaging 66 years, the heart will beat roughly 2.5 billion times.
Conduction system[edit]
Heartbeats are coordinated by an electrical conduction system, comprising:
- Sinoatrial node (SA node) – the heart’s natural pacemaker
- Atrioventricular node (AV node) – transmits impulses from atria to ventricles
- Bundle of His and Purkinje fibers – distribute electrical signals throughout ventricles, ensuring coordinated contraction
Clinical significance[edit]
The study and treatment of heart diseases and disorders are central to cardiology. Common cardiac conditions include:
- Coronary artery disease (blockage of heart arteries)
- Heart failure (insufficient pumping ability)
- Arrhythmias (abnormal heart rhythms)
- Cardiomyopathy (heart muscle disease)
- Valvular heart disease (valve dysfunctions)
Diagnostic methods include echocardiography, electrocardiography (ECG), cardiac MRI, and cardiac catheterization.
Historical perspectives[edit]
Throughout history, the heart has held significant cultural and symbolic importance, representing emotions, vitality, and life itself. The modern understanding of cardiac anatomy and physiology advanced greatly with discoveries such as William Harvey’s elucidation of blood circulation in the 17th century.
Gallery[edit]
-
 Heart
Heart -
 Heart
Heart -
 Heart
Heart -
 Heart
Heart -
 Heart
Heart





See also[edit]
External links[edit]
| Anatomy of the heart | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Physiology of the cardiovascular system | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|


Medical Disclaimer: WikiMD is for informational purposes only and is not a substitute for professional medical advice. Content may be inaccurate or outdated and should not be used for diagnosis or treatment. Always consult your healthcare provider for medical decisions. Verify information with trusted sources such as CDC.gov and NIH.gov. By using this site, you agree that WikiMD is not liable for any outcomes related to its content. See full disclaimer.
Credits:Most images are courtesy of Wikimedia commons, and templates, categories Wikipedia, licensed under CC BY SA or similar.
Translate page: - East Asian
中文,
日本,
한국어,
South Asian
हिन्दी,
தமிழ்,
తెలుగు,
Urdu,
ಕನ್ನಡ,
Southeast Asian
Indonesian,
Vietnamese,
Thai,
မြန်မာဘာသာ,
বাংলা
European
español,
Deutsch,
français,
Greek,
português do Brasil,
polski,
română,
русский,
Nederlands,
norsk,
svenska,
suomi,
Italian
Middle Eastern & African
عربى,
Turkish,
Persian,
Hebrew,
Afrikaans,
isiZulu,
Kiswahili,
Other
Bulgarian,
Hungarian,
Czech,
Swedish,
മലയാളം,
मराठी,
ਪੰਜਾਬੀ,
ગુજરાતી,
Portuguese,
Ukrainian
